
tags: researchblogging.org, Early Bird Project, Tree of Life, avian evolution, deep avian evolutionary relationships, avian phylogenomics, location cues, Shannon J. Hackett, Rebecca T. Kimball, Sushma Reddy
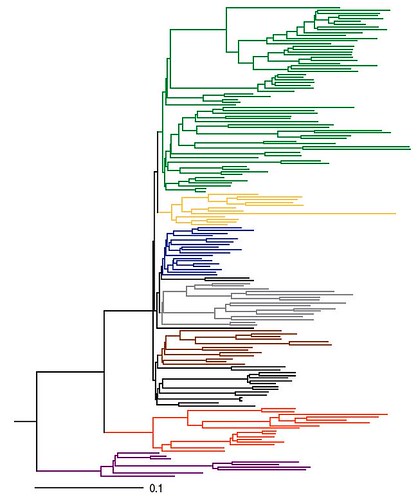
Basic topology of the evolutionary relationships between birds.
Maximum Likelihood (ML) phylogram reveals the short internodes at the base of Neoaves and highlights certain extreme examples of rate variation across avian lineages. The phylogenetic tree was rooted to crocodilian outgroups (not shown). Branch colors represent major clades supported in this study: land birds (green), charadriiforms (yellow), water birds (blue), core gruiforms and cuckoos (gray), apodiforms and caprimulgiforms (brown), galloanserae (orange), and paleognaths (purple). Scale bar indicates substitutions per site.
Image: SJ Hackett et al. DOI: 10.1126/science.1157704.
[larger view].
![]()
A fascinating paper was just published by some of my colleagues in the top-tier journal, Science, that analyzes the largest collection of DNA data ever assembled for birds. This analysis effectively redraws avian phylogeny, or family tree, thus shaking up our current understanding of the early, or "deep", evolutionary relationships of birds. For example, one of the most surprising findings of this analysis is that parrots and songbirds are "sister groups" -- each other's closest relatives!
And here's another surprise; falcons are the sister group to the parrots and songbirds. Further, the falcons (Falconidae) include the New World vultures -- but Falcons are not closely related to eagles, hawks and osprey (Accipitridae), as previously thought.
So why is avian taxonomy suddenly in such a state of upheaval? The precise evolutionary relationships between major groups of birds have long been contentious because they underwent an explosive radiation event sometime between 65 million and 100 million years ago. Nearly all of the major avian groups arose within just a few million years -- a very short period of evolutionary time. As a result, those groups of birds, such as parrots, doves and owls, that are united by distinct morphological characteristics seem to have appeared suddenly because there are few, or no, known evolutionary intermediates that provide clues to their deeper relationships with other avian groups.
These new findings are important because much of our scientific knowledge about reproductive biology, speciation, behavior, ecology, and natural history of animals is based on our studies of birds, so these data provide a new evolutionary framework that can be scientifically tested for decades to come.
This research, which was carried out over a period of five years by an international group of 18 scientists known as the Early Bird Assembling the Tree-of-Life Research Project, represents the most extensive genome-level analysis for any group of animals so far. Interestingly, the three lead authors of this study are female and more than half of "the Early Birds" are women.
To do this work, the Early Bird group examined 19 independent loci consisting of 32 kilobases of nuclear DNA sequence obtained from 169 avian species representing all major groups of birds alive today. These nuclear loci included introns (74%), exons (coding regions, or "genes") (23%), and untranslated regions (UTRs) (3%) across 15 different chromosomes (according to the chicken genome). These DNA sequence data were subjected to multiple statistal analyses that produced a robust phylogeny.
These analyses reveal two major findings: First, the classifications and conventional wisdom regarding the evolutionary relationships among many birds is wrong. Second, birds that have similar appearances or behaviors are not necessarily related to each other. According to these data;
- Birds adapted to different environments, such as terrestrial or ocean life, several times.
- Distinctive lifestyles evolved several times. For example, contrary to conventional thinking, the colorful, daytime hummingbirds are a specialized subgroup of the drably colored nocturnal or crepuscular nightjars; falcons are not closely related to hawks and eagles; flighted tinamous arose from the flightless rheas and ostriches; and tropicbirds are not closely related to pelicans and other waterbirds.
- Shorebirds are not a basal evolutionary group, which refutes the widely held view that shorebirds gave rise to all modern birds.
This phylogeny both confirms and challenges our current understanding of the relationships between the majority of bird groups as determined primarily on the basis of morphological analyses (figure 2);
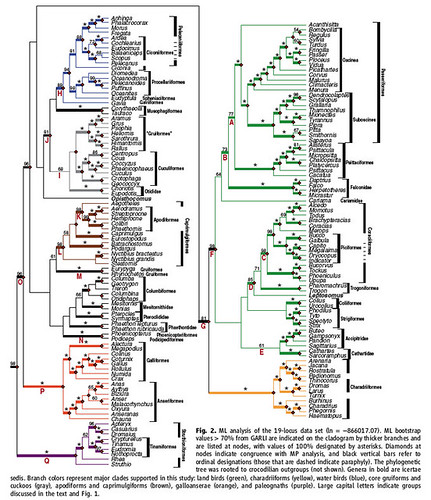
Figure 2: ML analysis of the 19 loci data set. ML bootstrap values > 70% from GARLI are indicated on the cladogram by thicketr branches and are listed at nodes, with values of 100% indicated by asterisks. Diamonds at nodes indicate congruence with MP analysis, and black vertical bars refer to ordinal designations (those that are dashed indicate paraphyly). The phylogenetic tree was rooted to crocodilian outgroups (not shown). Branch colors represent major clades supported in this study: land birds (green), charadriiforms (yellow), water birds (blue), core gruiforms and cuckoos (gray), apodiforms and caprimulgiforms (brown), galloanserae (orange), and paleognaths (purple).
Image: SJ Hackett et al. DOI: 10.1126/science.1157704 [larger view]
In a nutshell, this research confirms two previous taxonomic classification schemes that were based on morphological studies (also refer to Figure 4, below);
- Aves consist of two main groups: Paleognathae (ratites and tinamous) are separate from Neognathae (all other birds)
- Neognathae are split into two groups; the Galloanserae (ducks, chickens and their allies) and the Neoaves (other neognaths)
However, this research also challenges many of our current taxonomic schemes (Figure 2, see also Figure 4, below);
- The largest clade within Neoaves consists of the landbirds (green): Passeriformes (songbirds), Piciformes (woodpeckers and allies), Falconiformes (falcons), Strigiformes (owls), Coraciiformes (kingfishers, hornbills, rollers and allies), Psittaciformes (parrots), Coliiformes (mousebirds) and Trogoniformes (Trogons). Landbird surprises include;
- Parrots and songbirds are sister groups.
- Falcons are the sister group to parrots and songbirds.
- Falcons (Falconiformes) are a distinct clade from the eagles, hawks and osprey (Accipitridae).
- New World vultures (Cathartidae) do not belong in the storks and allies (Ciconiiformes), but should instead be included with the falcons (Falconiformes).
- Woodpeckers (Piciformes) are a specialized grouping within the kingfishers, hornbills, rollers and allies clade (Coraciiformes).
- Seriemas (Cariamidae) are sister to the falcons not the Gruiformes.
- The sister group to the landbird clade are the waterbird clades: Charadriiformes (shorebirds, gulls, and alcids) (yellow), and the Pelecaniformes (totipalmate birds), Ciconiformes (storks, herons, bitterns and allies), Procellariiformes (tube-noses), Sphenisciformes (penguins), and Gaviiformes (loons) (all blue). Additionally;
- Buttonquail, Turnix species, belong within Charadriformes.
- Pelicans (Pelecaniformes) and storks, herons, bitterns and allies (Ciconiformes) form one clade.
- The tropicbirds (Phaethontidae), which were previously classified within the pelicans (Pelecaniformes), are now excluded from the newly identified Pelecaniformes-Ciconiformes clade.
- Some other revelations include;
- Hummingbirds and swifts (Apodiformes) share a common ancestor with the nightjars (Caprimulgiformes).
- Sunbitterns (Eurypygidae), which are neotropical species, and the mysterious New Caledonian Kagu, Rhynochetos jubatus, are sister taxa that are not found within Gruiformes.
- Grebes (Podicipediformes) and flamingos (Phoenicopteriformes) are sister taxa.
- Tinamous (Tinamiformes), which are flighted, arose within the flightless Struthioniformes (rheas, ostriches, cassowaries, emus and kiwis).
Yet, despite the large sample size and the rigor of the analyses, there's still a few avian mysteries whose relationships remain unresolved;
- Hoatzin, Opisthocomus hoazin -- just what is this creature??
- Pigeons and doves (Columbiformes).
- Sandgrouse (Pteroclididae).
- Tropicbirds (Phaethontidae).
- Are Caprimulgiformes (with owlet-nightjars removed) monophyletic or paraphyletic?
- and the appropriately named mesites (Mesitornithidae), is a family of birds of uncertain affinities (this is sometimes known as the "taxonomic garbage can"). But maybe, since the three mesites species are all endangered anyway, perhaps they all will be wiped out forever, so we no longer have to worry about how they fit into the evolutionary tree for birds?
These new data correspond for only six avian orders when compared to two studies that were published earlier; Sibley and Monroe's DNA-DNA hybridization study (1990) and Livesey and Zusi's comparative anatomy study (2007) (figure 4);
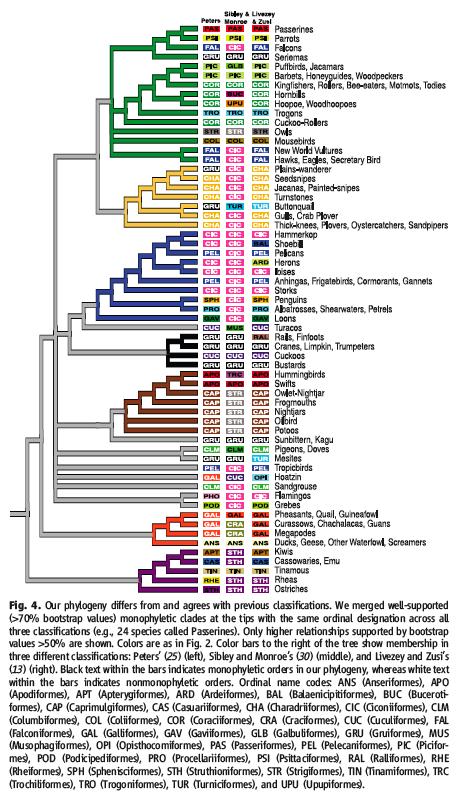
Image: SJ Hackett et al. DOI: 10.1126/science.1157704.
In light of these new data, comparative studies of birds will benefit from increased rigor and will yield greater insights into the process of evolution and speciation. This research also affect publishers and birders because biology textbooks and birdwatching field guides will have to be rewritten.
Notes on the phylogeny and questions for the Early Birds:
- What happens if you remove Leptosomus (Clade D)? Does this clade resolve with stronger support without it included in the analysis?
- I am surprised that the connection between Passeriformes and Psittaciformes and the connection between Passeriformes-Psittaciformes and Falconiformes are being taken so seriously when they have such low bootstrap support [but see Ericson et al. (2006) DOI: 10.1098/rsbl.2006.0523]. What gives? What would happen to the analysis if the Falconiformes were removed? How much rearranging of the Passeriformes, Psittaciformes and Charadriiformes within the tree itself would occur?
- Which gene(s) place seriemas within Falconiformes?
- Accipitridae show abberant behavior in molecular phylogenies, and everyone agrees that this lineage is old, and the bootstrap support for this clade is not strong.
- The fossil record strongly suggests that the Charadriiformes should precede Ciconiiformes along with a few other avian orders, and the fossil record for Turnix suggests it predates gulls and alcids. And wow, I thought sandgrouse were located within this clade?? Which gene(s) relocated them?
- Have you noticed that, if Charadriiformes is removed, the support for much of the tree is lost. What gives here?
- There is surprisingly poor taxon sampling for the non-Passeriodea Passerida.
- I am disappointed by the really poor bootstrap support for the Columbiformes node .. knowing the placement for this order would really help clarify our understanding of avian relationships in general.
- The placement of Tinamous within the ratites instead of being basal to them suggests either that flight was lost and gained several times in the evolutionary history of birds or that flight was lost independently many times, but only within the ratites. Weird!
Source:
Hackett, S.J., Kimball, R.T., Reddy, S., Bowie, R.C., Braun, E.L., Braun, M.J., Chojnowski, J.L., Cox, W.A., Han, K., Harshman, J., Huddleston, C.J., Marks, B.D., Miglia, K.J., Moore, W.S., Sheldon, F.H., Steadman, D.W., Witt, C.C., Yuri, T. (2008). A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science, 320(5884), 1763-1768. DOI: 10.1126/science.1157704.
Also see:
Ericson, P.G., Anderson, C.L., Britton, T., Elzanowski, A., Johansson, U.S., Källersjö, M., Ohlson, J.I., Parsons, T.J., Zuccon, D., Mayr, G. (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters, 2(4), 543-547. DOI: 10.1098/rsbl.2006.0523.
I thought the early bird got the worm?
(Sorry, couldn't resist!)
HOLY SMOKES! Bird phylogeny will never be the same. I saw this post and skimmed it fast, thinking oh great, another phylogeny which changes up everything, how is this one any different, but then I saw they used 19 LOCI! Wow. I am in awe. Good work, Hackett et al. Thanks for bringing this to my attention. I think I'm going to spend half the night poring over it.
~ Nick
Thanks for this thorough report on the Science paper and for including the cladograms. It will be interesting to see what the various checklist committees do with the results.
"The phylogenetic tree was rooted to crocodilian outgroups (not shown)"?????
I thought that it was widely accepted that birds are the last dinosaurs. Or are dinosaurs "crocodilian outgroups?"
No, crocodilians are dinosaur outgroups. Crocodilians are, however, the closest available outgroup for molecular analysis of birds, seeing as everything closer has been extinct for too long for much molecular data to be available.
How does the "tastes like chicken" trait map onto the tree?
Or rather, flight was lost three times independently within paleognaths: once in ostriches, once in rheas, and once in kiwis + emus + cassowaries... or rather twice within the latter clade, because imagining a kiwi rafting from Australia to New Zealand is a bit hard.
But the position of the ostrich in the tree is suspect anyway -- it looks like long-branch attraction. Look at Fig. 3, the one on top of this blog post: the ostrich is the purple branch at the very bottom. That the ostrich has a faster rate of molecular evolution than usual has long been known.
There might even be long-branch repulsion between the ostrich and the remarkably long branch at the base of the tinamous.
Preliminary conclusions were presented at the paper session of the Minnesota Ornithologists Union that I attended this Spring. As the conclusions were introduced, the sounds of those in the auditorium murmuring around me fell away, and visually everything became blurred out except the presenter and the phylogeny on the screen as my perceptions of how evolutionary processes must have produced these birds shifted, blew apart and refused to settle back down into coherence. Thank you so much for your post on this subject.
You said:
Further, the falcons (Falconidae) include the New World vultures -- but they are not closely related to eagles, hawks and osprey, as previously thought.
..but later explain that New World vultures are actually in Accipitridae after all.
OOPS! thanks for spotting that error.
1. I don't see how a flightless bird can re-evolve flight. Impossible I would have thought.
2. How can you tell from DNA that the common ancestor of the hummingbirds and nightjars was nocturnal?
You wrote:
Did you actually mean that Seriemas are sister to the grouping of falcons, parrots, and passerines? That's how it appeared to me.
Also, how does one explain, from a biogeographical approach, the locations of the sunbittern and kagu if they are sister to one another... Gondwanaland was split far too early for the ancestor of this clade to have occupied the entire continent, right? Does one invoke island hoping? The new world tropics are a bit removed form New Caledonia.
Excellent writeup. Thanks so much for including the figures!
Not from DNA, but from the tree: either a nocturnal lifestyle evolved several times independently in this clade, or it evolved once and reversed once. The principle of parsimony favors the latter hypothesis.
-------------
New Caledonia and South America were connected via New Zealand and Antarctica till 88 million years ago, which is way too early, but the sunbittern can fly, and the connection between South America and Antarctica only broke 37 or so million years ago...
And aren't there fossil sunbitterns in the early Eocene of North America?
I should have mentioned that the New World tropics weren't far removed from Antarctica 40, let alone 55, million years ago. Paratropical rainforest ( = with a short dry season) extended into Antarctica even.
Fantabulous! A feather in your cap for writing it up for us.
'How can you tell from DNA that the common ancestor of the hummingbirds and nightjars was nocturnal?'
'Not from DNA, but from the tree: either a nocturnal lifestyle evolved several times independently in this clade, or it evolved once and reversed once. The principle of parsimony favors the latter hypothesis.'
Why can't it have evolved once, after the split between hummingbirds and nightjars?
We know from both molecular and morphological data that Apodiformes (swifts and hummingbirds), which are diurnal, are nested within Caprimulgiformes (nightjars and allies) rather than being their sister group. So therefore if nocturnality (if that's a word) evolved in nightjars after they split from Apodiformes, it would have had to have evolved independently in owlet-nightjars, oilbirds and frogmouths - owlet-nightjars are more closely related to Apodiformes than nightjars, while oilbirds and frogmouths are less closely related to Apodiformes. As David pointed out, there would have been less changes in lifestyle involved if diurnality is a reversal in Apodiformes than if it evolved separately in each of the caprimulgiform families.
Common Swifts here in Europe are known to sleep on the wing, only landing normally on rare occaisions outside the breeding seson, and I presume other species are similar. Since nightjars and swifts both catch flying insects by aerial "filter feeding" for want of a better term, a switch from feeding on nocturnal to feeding on diurnal insects would be all that is nedded to turn a nightjar into a proto-swift
The way I learnt it was, Ratites develop the flight muscles later than other birds, so it's easier for them to ditch flight if there's no reason to keep it, and so an ancestral group of flighted ratites spread out and then lost flight individually to make use of ground resources. In response to predation, some groups grew really big and developed a nasty kick (Ostriches, Emus, Rheas, Cassowaries and presumably Moas) and one group shrank and became nocturnal (Kiwis), a strategy which also worked for other birds which lost flight (Kakapo). Tinamous, on the other hand, for some reason never lost flight.
As for falcons and parrots being related, seeing as Keas are virtually birds of prey, it's not too hard to imagine. Songbirds, not so much.
"...falcons (Falconidae) include the New World vultures -- but they are not closely related to eagles, hawks and osprey (Accipitridae), as previously thought."
In fact that whole "land-birds" clade can be seen as a "birds of prey" clade with non "flesh"-eating reversals sprinkled between the falcons, hawks, and new world vults, so Falconidae & Accipitridae are characteristic occupiers of the original broad niche of a reasonably old clade. Carnivores spawning non-carnivorous offshoots is a recurring theme amongst vertebrates.
I love the idea of parrots having inherited their hooked (upper) beaks from birds of prey - a day-dream I have long treasured. And although it isn't quite what their cladogram suggests, I've always thought sapsuckers (a branch of woodpeckers) look very similar to orioles or corvids e.g nutcrackers. You say there is low bootstrap support in that part, so perhaps I can hope!
The survival across the end of the Cretaceous of a number of paleognath lines (if this is not an artifact) justifies the authors' claim to have refuted (again) the view that shorebirds gave rise to all modern birds. However the explosion suggested by their fig.3 (presumably the start of the Palaeocene) leaves the possibility that outside the paleognaths, one "duck" and one other bird species survived the K to leave descendants today. That other bird led to the greatest explosion, and I'm guessing that means it was a water bird, best able to travel widely and access universal habitats in the scorched new world. So perhaps 99% of modern birds were descended from an early duck and (the vast majority) from that presumed shorebird.
The tinamou would not have had flightless ancestors more recent than the Jurassic. Regaining flight after losing it probably never occurred; the paleognaths seem to have had a tendency to lose flight.
Thanks for your posting.
I studied the White-breasted Mesite and have worked on the conservation of the three (threatened) species in this very odd family. THey have always turned out incertae sedis in this kind of analysis. Surely that suggests that they are potentially a very interesting taxon worthy of more detailed study? Is anybody digging deeper on their taxonomy?
Will we ever have an holistic and comprensive work related to Phylogeny? Genetics are the key? Some people talk like it is. Is morphology the key?
What I know is that tomorrow (literally) this could change because some other group of scientist use another set or part of the bird's genome.
So..I'll read it, and wait for someone to bring to light a paper having a comprehensive work including, ecology, behaviour, genetics and morphology (and whatever you want to add). It is not an UTOPY.
By the way does someone have an ornithology book (pdf,zip) or related to birds to share (rapidshare, megaupload, etc.)?
Cheers
Lucas
I've had some time to digest this work and write about it:
http://slybird.blogspot.com/2008/07/avian-relationships-what-do-we-know…
As far as your questions...
I don't think sandgrouse (Pteroclidae) were ever considered Charadriiformes? I think they are most frequently considered related to or within Columbiformes, but I could be wrong on that count.
Do you have any references for Accipitridae's aberrant behavior in phylogenies?
As there is little doubt that Passerines are monophyletic, why would they try to sample extensively in it? It will take a tree even bigger than Hackett et al. just to work out Passerine relationships with satisfaction.
I echo your questions about the different possible trees. I really wish they had provided alternative trees for some of the things you suggest, and for taking b-fib out to remove the Metaves clade.
~ Nick
Hi. Enjoyed your blog entry, and I thought I would try answering the questions you had for Early Bird.
1. What happens if you remove Leptosomus? We don't know. Just doing the analyses we actually did took months of supercomputer time. There are obviously many analyses it would be interesting to do with our data that we haven't got to. Taxon jackknifings are one huge class of such analyses. Same answer to other questions you asked about what happens if you remove various taxa.
2. Why focus on passerine-parrot and passserine-parrot-falcon nodes when they don't have great support? Simply because they're so interesting. And cool. And there is a bit of support. The only single-gene support comes from EEF2 and SOMA, but notice that if you jackknife each of those genes, the nodes are still there. We don't know what happens if you jackknife both EEF2 and SOMA; multi-gene jackknifing is another example of analyses it would be good to do that we haven't. As for strong support in Ericson et al. for this group, note that the support numbers there were Bayesian posteriors, which have been shown in several papers (and in my experience) to be highly inflated measures of confidence. As expected, the support from 5 genes is not greater than the support from 19. (Notice also that 3 genes are shared between data sets.)
3. Which genes palce seriemas within falconiforms? None, to my knowledge. The phrase "near other raptorial groups" refers to their being inside "land birds", and near the falcons.
4. Low bootstrap for Accipitridae. Actually, the bootstrap for Accipitridae is 100; you probably mean Accipitriformes, the group including accipitrids, ospreys, secretarybird, and NW vultures. Only the last is ambiguously attached to the rest, and it's a wonder that we got that much, if you ask me.
5. Fossil record, sandgrouse. I'm not sure the fossil record of birds is good enough to show what orders precede others, and one can always distinguish between stem groups (which don't necessarily have all the diagnostic characters of the group) and crown groups (which do). So in fact Charadriiformes (crown group) could precede Ciconiiformes (crown group) even if Ciconiiformes (stem group) diverged from other orders before Charadriiformes (stem group). Turnix ditto, though in fact its position in our tree is outside the node connecting gulls and alcids, so there should be no problem even if we ignore the stem/crown distinction. As for sandgrouse, they have bounced back and forth between charadriiforms, columbiforms, and their own private order in traditional classifications. I don't know of any molecular data that resolve their position.
6. If charadriiforms are removed, much support is lost. Really? Of course we haven't done this analysis or any other taxon jackknifing. Did you try it? Could you elaborate?
7. Poor taxon sampling for non-passeroid passerids. What you really mean is that passeroids are shockingly oversampled. The reason for that is that we chose on purpose to sample each species for which there was a sequenced mitochondrial genome at the time, and that included, for some reason, 3 passeroids. Our intention was not to resolve phylogeny within passeriforms (whole nother grant there) but to sample as evenly as possible through all of bird diversity, as estimated from SIbley & Ahlquist distances. Passerines as a groups are actually somewhat oversampled by this criterion.
8. Disappointment at poor resolution of some nodes. Me too. "Metaves" seems to consist entirely of difficult-to-connect clades. I would like to have twice the data. But I'm also pleased with what we got.
9. Tinamou placement is weird. You bet. But look at how many of the individual gene analyses support it. It's as weird as grebes plus flamingos, and as easy to find with nuclear genes. By the way, this is clearly not long branch attraction, as one comment had it. We've done analyses to check that. If, for example, we simulate LBA with a randomized outgroup, the root actually attaches either at the tinamous (the traditional position) or within them. And relative rate tests show that the ostrich is not any faster-evolving for our genes than are other ratites. Tinamous, now, they're bizarrely fast for everything. No idea what's going on there, but it's a genome-wide phenomenon.
I think Hoatzin line probably developed long before, or at the same time as, the rise of modern birds. Kind of like how the Tuatara line predated or coincided with the development of lizards. Even though it sure looks like a lizard, it's not one.
Several independent losses of flight in ratites are not a surprise, but...
That's really surprising, because several previous papers reported just that -- I don't know for which genes, but I think the complete mito genome was among them.
Interesting.
--------------------------
There is no reason whatsoever to think so. It's an ordinary modern bird. All that's going on is that its finger development is delayed -- one mutation.
Though... if the root of Neornithes as a whole attaches to the tinamous, that's not the traditional position, although it's close. Is that what you meant? The traditional position for that is between paleo- and neognaths, both thus being monophyletic.
I am definitely not an expert here, but I thought it was conventional wisdom that the Hummingbirds were most closely related to the Nightjars.
The first time I saw a hummingbird in a natural setting, as opposed to a garden or a hummingbird feeder, it was catching flying insects. I said I thought that was strange, and the resident expert of our party told me that hummingbirds ate lots of insects, and were descended from Nighthawks. This was in either 1989 or 1990.
Couple of responses:
No, tinamous were not (when adding a random outgroup) the root of Neornithes, but of Palaeognathae; i.e., the traditional position.
Hoatzins belong to Neoaves, and thus can't predate all other birds, whatever their relationships within Neoaves, as yet undetermined.
Conventional wisdom has been that hummingbirds were most closely related to the swifts, and nothing about that has changed. It has often been thought that the swift/hummingbird clade was most closely related to the nightjars (not really -- nightjars are a family, Caprimulgidae, while the conventional relationship was to the entire order Caprimulgiformes, including several families). What's interesting about our phylogeny (and several other recent ones) is that Caprimulgiformes is paraphyletic to the swift/hummingbird clade, meaning that some caprimulgiforms are more closely related to hummingbirds than to other caprimulgiforms. This suggests that a nightjar-like lifestyle is ancestral to the hummingbird lifestyle. Without paraphyly, there is no evidence for this, so whoever told you this in the '89/90 was prescient in the absence of data. Hummingbirds eat some insects, especially when feeding nestlings (nectar isn't a very good food for baby birds).
In Comment 14 David M asked about fossil sunbitterns in from the Eocene of Wyoming. I just did a pretty comprehensive report on the fossil birds of Wyoming. Here's what I know, there are a few fossils of "primitive Gruiformes" that are attributed to being closely related to sunbitterns and kagus. (there they are again in the same sentence). As far as I know, the work is not published scientifically, only in semi-popular press as a preliminary assessment type of identification. (I might have seen it in Feduccia's book, but don't quote me).
Meanwhile, as a birdwatcher and paleontologist (of sorts) I'm always blown away by the stuff I see coming out of the DNA-ornithlogy camps.
wanna bet that this too will change???
Ah, thanks. Will be interesting to see where they fall once a phylogenetic analysis is done <crickets chirping>.
Sure, but not much anymore. It's largely congruent with the last few analyses, and the unexpected result of ratite polyphyly was confirmed by a recent PNAS paper (came out last week or so) that dealt specifically with this issue.
In the paper:
Wink, M. 2000. Advances in DNA studies of diurnal and nocturnal raptors. In Chancellor, R.D. & Meyberg, B.-U. (Eds.), RAPTORS AT RISK, 831-844. WWGBP, Berlin and Hancock House, Surrey (Canada).
this "old" paper did suggest Falconiformes (Falconidae) and Accipritriformes (Accipitridae) as separate and even Elanus kites as Elaniformes. I also am interested in whether Northern Harriers are Circus cyanea hudsonia or Circus hudsonia (hudsonius?). The AOU I believe (?) says the former.
I emailed The Chicago Field Museum and they said that convergence between Falconiformes and Accipitriformes is on the list of things to investigate.
Another ornithologist lamented that a lot of systematic work is done perhaps at the expense of natural history, which is still wide open. Let me say that birders can still publish in regional journals original behavior, biological, and ecological observations.
you all seem quite bright,...
No offense ment,...
I was looking up the cladistics of the ratites... never knew that there was so much discrepancy.
Have we ever thought of a background in plate tectonics, and a similar background in botony?
These questions as to cladistics, shouldn't be such a problem,... certain plants only grow, in certain areas, across the globe, within this time frame, but have occured in different areas, when we were Pangea, that's a given.
But, given that plants don't expand world wide, unless given help by humans, and less by migritory birds,...it's just a thought.
I just seem to think, even given the recent discovories, of feathered dinosaurs, and the acceptance of dinosaurs evolving into birds,... it shouldn't be that hard to see the relationship of birds to each other.
Just a thought,...
Don't be afraid to respond,... I've been called stupid before,...
Kindda used to it.
But, I feel that if we knew more about the ecology, given with what we know of birds to date, and their specialities to plants dictates, ...as to what we've learned so far, should be a better place to start, to find a resolution to their cladistics.
I didn't really understand why the ornithologist got so excited about the Great Tinamou we ran across in the field. Now I think I get it ;)
I'm really far from an expert, but from the little I've seen of Tinamous they posses the ability to "fly" only in a rather generous meaning of the term. I can easily imagine paleognaths as obligatory ground birds with a high propensity to loose whatever little flight ability they have. Loss of function is generally easy evolutionarily speaking.
On the other hand, regaining a previously lost function can also be relatively easy depending on the physiological mechanisms and number of epistatic adaptations build on it. There may be some really interesting EvoDevo angles in there... someone page PZ.